
Autor: Pedro Pablo Fernández
Interacción entre sistemas energéticos
La contracción muscular y, por tanto, todo ejercicio físico dependen de la ruptura del ATP (Adenosine TriPhosphate o Adenosín Trifosfato) y su correspondiente liberación de energía.
La cantidad total de ATP almacenada en las células es muy pequeña (aproximadamente 8 mmol/kg de peso seco de músculo). Por ello las células tienen que utilizar otros mecanismos para aportar ATP que mantenga la actividad muscular y para almacenar energía en moléculas más complejas.
Como se ha dicho, la concentración de ATP inicial es reducida, pero con procesos de utilización de ADP, AMP y Pi, se puede seguir haciendo frente a elevadas demandas de energía para el funcionamiento de la célula.
Es importante destacar que el organismo (principalmente el tejido muscular), ante requerimientos de ATP, es capaz de variar su nivel de producción metabólica en dependencia de las demandas energéticas.
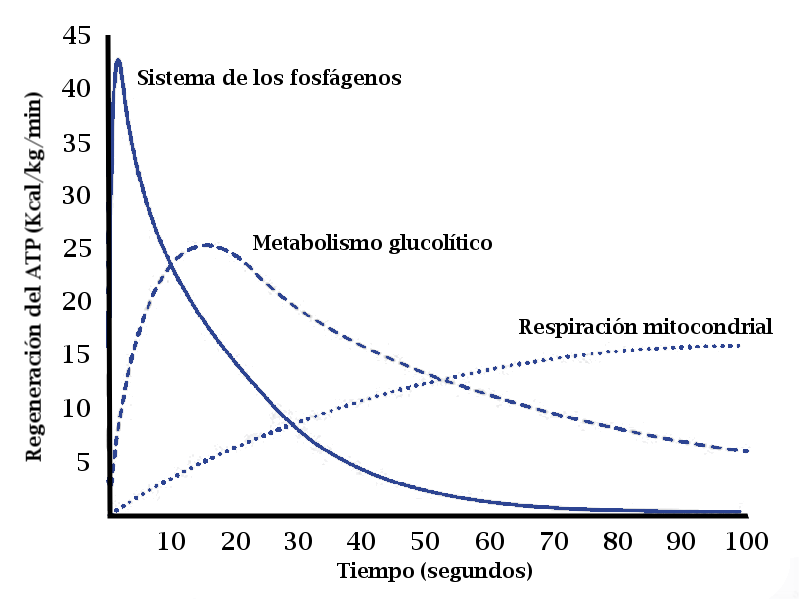
Hay tres sistemas responsables de la resíntesis de ATP:
- El sistema de los fosfágenos.
- El metabolismo glucolítico.
- La respiración (oxidación) mitocondrial.
Los sistemas energéticos responden en función de la cantidad y velocidad de ATP que hay que resintetizar en la unidad de tiempo, es decir, de la intensidad del ejercicio.
Desde un punto de vista más bioquímico, las vías de resíntesis de ATP serían:
- La ruptura/hidrólisis de CrP/Fosfocreatina (incluso de ADP).
- La fosforilación de glucosa no dependiente del oxígeno: glucolisis anaeróbica.
- La fosforilación oxidativa de glucosa: glucolisis aeróbica.
- La lipolítica: fosforilación oxidativa de lípidos (grasas).
- Y, en algunos casos, la proteolítica: la fosforilación oxidativa de proteínas.
Metabolismo anaeróbico
- El sistema de los fosfágenos.
- El sistema glucolítico.
El metabolismo anaeróbico (no mitocondrial) es capaz de actuar de inmediato ante las demandas de energía, de ejercicios de intensidad alta. Pero tiene una capacidad limitada, por lo que si el ejercicio continúa, o bien hay que parar por fatiga o hay que reducir su intensidad hasta un nivel en el que la energía pueda obtenerse mediante el metabolismo aeróbico (oxidación mitocondrial).
Teóricamente, se sabe que en los deportes que requieren contracciones musculares intensas, rápidas y repetidas (velocidad, saltos, fútbol, tenis, etc.) la fosfocreatina (CrP) es la única responsable de la regeneración y aporte de ATP durante los primeros 8”-10”. La hidrolisis de la fosfocreatina no depende de la disponibilidad de oxígeno, ni necesita reacciones químicas complejas, para liberar energía con objeto de resintetizar ATP.
En la cinética de la resíntesis de la CrP/fosfocreatina, se observa mayor rapidez del proceso, cuando hay un porcentaje menor de velocidad de vaciado (ejercicio submáximo) y cuando las fibras implicadas son de tendencia Ft (rápidas, tipo IIb/FTg y IIa/FTo).
Sin embargo, se ha demostrado que la glucolisis se activa muy rápidamente durante el ejercicio intenso y regenera ATP antes de lo que se creía hasta ahora.
Si la fosfocreatina, solo se repone parcialmente durante la recuperación, puede comprometer el rendimiento deportivo, principalmente en aquellas especialidades donde hay que repetir las acciones de intensidad máxima (rugby, fútbol, etc. Aunque no afecta al 400m).
La glucolisis (anaeróbica y aeróbica)
Es un error entender la glucolisis como un proceso anaeróbico, ya que, efectivamente tiene una fase no dependiente del oxígeno y otra que sí depende de él (si el ejercicio es intenso, como es el caso de los 400m, se producirá lactato independientemente de la oxigenación).
Hay que recordar que el glucógeno se utiliza tanto en el mecanismo anaeróbico, como en el aeróbico.
Cuando el ejercicio tiene una intensidad submáxima (o su primera fase es submáxima, caso del 400m) y continúa más allá de los 10”, la energía para regenerar ATP proviene de la glucosa sanguínea y el glucógeno muscular.
La vía glucolítica se activa muy rápido cuando se realiza ejercicio intenso (prácticamente desde el inicio del ejercicio), aunque a diferencia de la hidrólisis de la CrP, que alcanza su máximo rendimiento instantáneamente, la producción de ATP mediante la glucolisis, no alcanza velocidades altas hasta pasados 10”-15”.
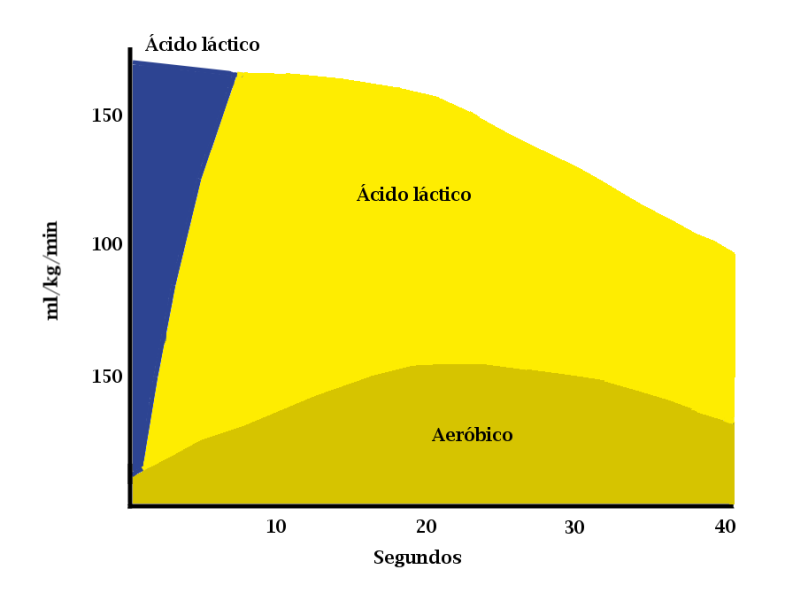
“Se ha comprobado que en un esfuerzo máximo de 30”, los fosfágenos aportan el 23% de la energía, la glucolisis el 44% y el 28%, la respiración mitocondrial”.
En situaciones mayoritariamente anaeróbicas (caso del 400m), se obtiene poca pero rápida energía del glucógeno y, en proporción, se gasta mucho más que en la situación aeróbica.
Esta actuación tan inmediata de los procesos energéticos relacionados con los carbohidratos, justo después del inicio del ejercicio, se debe a la producción de AMP y al aumento intramuscular de Ca y Pi y también a la entrada de glucosa en el músculo activo.
La cantidad total de glucosa que se puede almacenar en forma de glucógeno es relativamente pequeña, hallándose principalmente en el músculo (adulto de 70 kg, 300/600 gr), el hígado (de 0 a 90 gr) y en el líquido extracelular (de 8 a 11 gr), dependiendo estas concentraciones de la dieta y la adaptación por el entrenamiento.
Esta vía produce lactato en los músculos activos y su liberación se produce enviándolo:
- A los capilares que rodean la fibra muscular, para ser transportado por la sangre al hígado, donde puede ser convertido en glucógeno o utilizado como fuente de energía por otros órganos (corazón, etc.) y
- a las fibras musculares cercanas, en las que puede usarse como sustrato energético.
“La producción de lactato en el músculo durante el ejercicio intenso es beneficiosa para eliminar piruvato, mantener los niveles altos de glucolisis y regenerar moléculas de NAD+, que servirán para el funcionamiento de la propia glucolisis”.
(Robergs y cols., 2004)
“Otro efecto beneficioso de la producción de lactato sería el hecho de que en la reacción de la LDH (LactatoDesHidrogenasa) se utilizan dos protones, con lo que se logra un efecto de tamponamiento de protones. Así pues, el lactato contribuye al reciclaje de protones liberados por la glucolisis y en la hidrólisis de ATP durante el ejercicio muscular”.
(N. Terrados, 2013)
La producción aumentada de lactato no es la causa de la acidez intramuscular durante el ejercicio intenso.
La acidosis (que es para muchos autores la causa de cierto tipo de fatiga) ha sido considerada como la consecuencia del aumento de la producción de ácido láctico muscular, que liberaba protones y formaba lactato.
Cuando la producción de lactato superaba la capacidad tampón muscular, el exceso de protones producía una disminución del pH de la célula muscular; lo que ha sido durante 80 años la explicación fisiológica de la llamada “acidosis láctica”.
En la actualidad, algunos autores (Robergs y cols. 2004) piensan que no hay evidencias de que la producción de lactato produzca acidosis muscular, sino todo lo contrario: “El lactato retrasa la acidosis muscular”.
Sólo cuando un ejercicio aumenta su intensidad por encima del estado estable y se necesita obtener mucho más ATP del sistema de los fosfágenos y de la glucolisis es cuando el ATP aportado por esas fuentes extra-mitocondriales aumenta la liberación de protones y causa la acidosis muscular.
La producción de lactato aumenta en esas condiciones metabólicas por acción de la LDH, para prevenir un aumento y acumulación de piruvato y para aportar el NAD+ que se necesita en la segunda fase de la glucolisis.
Este aumento de lactato coincide con la acidosis muscular y sigue siendo un buen indicador indirecto de las condiciones metabólicas del músculo.
Si el músculo no produjera lactato, la acidosis muscular y la fatiga se presentarían antes y afectarían al rendimiento.
“Los niveles de transportadores de Lactato aumentan con el entrenamiento”.
Al disminuir la potencia demandada (agotamiento del proceso anaeróbico), la energía tiene que limitarse a la que pueda producir el metabolismo aeróbico (la potencia demandada disminuye en relación con las limitaciones de las diversas fuentes de energía).
El metabolismo aeróbico responde a las demandas energéticas también más rápido de lo que se pensaba hace unos años, siendo evidente en la actualidad su papel en el trabajo de alta intensidad.
Así, en esfuerzos de alta intensidad de 75”, se obtendría la misma energía del sistema aeróbico que del anaeróbico. Evidentemente, el origen de la energía, según la vía solicitada, define con claridad la gráfica del rendimiento (en el caso del 400m, tanto en tiempos parciales como en metros por segundo). En la “gráfica del rendimiento” del 400m establecida por tomas de tiempos cada 50m, las zonas de los fosfatos, la glucolisis anaeróbica y la glucolisis aeróbica se definen con claridad y un alto grado de paralelismo, al margen del nivel del atleta. Una línea con un gran y breve crecimiento de inicio se define progresivamente descendente, a partir de la toma entre el 100m y el 150m, indicando claramente lo expuesto.
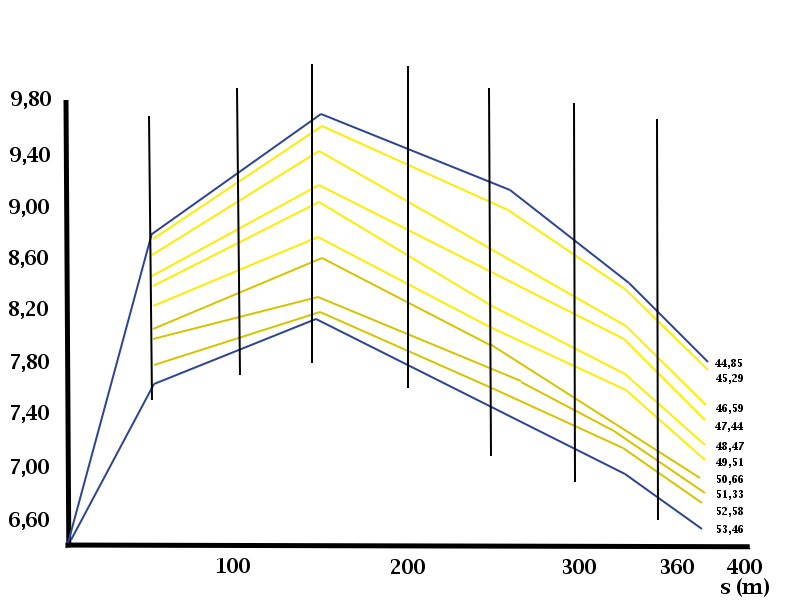
| Hombres | Mujeres | ||
| 400m | n | 400m | n |
| 44,85 | 5 | 50,66 | 19 |
| 45,29 | 7 | 51,33 | 24 |
| 46,59 | 63 | 52,58 | 35 |
| 47,44 | 46 | 53,46 | 35 |
| 48,47 | 43 | ||
| 49,51 | 18 |
| Fracción (m) | Tiempo (segundos) | Velocidad media (km/segundo) | Potencia media |
| 0-100 | 11,10 | 32,432 (-2,7%) | 180,59 (+12,0%) |
| 100-200 | 10,12 | 35,573 (+6,7%) | 179,74 (+11,5%) |
| 200-300 | 10,44 | 34,483 (+3,4%) | 160,91 (-0,2%) |
| 300-400 | 11,52 | 31,250 (-6,3%) | 126,56 (-21,5%) |
| 0-400 | 43,18 | 33,349 | 161,27 |
Aplicación práctica en el 400m
Como consecuencia de la confirmación, respecto a la anticipación y superposición de las vías energéticas, en lo que podría definirse como apoyo y resíntesis de aporte de energía ante demandas breves e intensas:
- La vía glucolítica anaeróbica apoya desde inicio a la vía de los fosfatos, llegando, en esfuerzos submáximos, a prolongar el aporte de ATP de primera instancia hasta los 15”-20”.
- La vía glucolítica aeróbica apoya a la anaeróbica con un protagonismo manifiesto en los porcentajes detectados, que se indican en estas consideraciones.
Nosotros, los entrenadores, deberíamos centrar nuestra atención en diseñar programas de desarrollo y mejora del rendimiento en dos direcciones principalmente:
- Adaptación al necesario control y desarrollo de la velocidad submáxima inicial (reparto óptimo del esfuerzo) que nos permita prolongar al máximo la fase de protagonismo de los fosfágenos (120 m >> 150 m >> 180 m >>… ) gracias a la resíntesis de ATP, desde el inicio, prolongando los recursos de la CrF mediante el “apoyo” de la glucolisis anaeróbica (esta alcanza su mayor velocidad a los 10”-15”). Se mejora por adaptación metabólica.
- Ante la muy importante caída del rendimiento en los últimos 80-100 m (protagonismo del sistema glucolítico anaeróbico/aeróbico), parece adecuado tratar de reducirla en lo posible, mejorando por adaptación metabólica la potencia de este último proceso energético y mejorando también el reparto del esfuerzo, de tal forma que permita un último 100 m en un porcentaje próximo al 26-26,50% del tiempo total previsto.
En relación con el primer punto, parece adecuado plantear unidades específicas de adaptación:
- El entrenamiento continuado y progresivo, de carácter submáximo, de series de 120/180 m (16”-20”), con el fin de estimular y favorecer una positiva adaptación del proceso de apoyo inmediato al consumo inicial de la CrP.
En la práctica, con atletas de 3-4 años de experiencia, esto supone poder realizar unidades de resistencia a la velocidad (prolongada), por ejemplo: 3 x 3 x 150 m con micro y macro recuperación de 5’ y 10’ sin apenas acúmulo de lactato (6-8 mmol) a los 30’-45’ de terminar la unidad, permitiendo, tanto después de dicha unidad como al día siguiente, la realización de ciertos tipos de trabajo compatibles con lo llevado a cabo.
Está claro que esta primera tendencia conllevará paralelamente un cierto desarrollo de la velocidad máxima del atleta, planteando las unidades en condiciones previas favorables y sobre recorridos y registros en velocidad lanzada, tramos de 50-60 m con 20-30 m previos, primando la frecuencia, soltura y decontracción.
Ambas unidades deben tener cabida en dependencia del planteamiento de los periodos y/o mesociclos, al término del denominado general, elaborando un programa que abarque el especial y competitivo.
Asimismo, la mejora del segundo aspecto requerirá como complemento previo aconsejable la utilización en el periodo general de unidades de capacidad láctica / potencia aeróbica glucolítica (% bajos: 80-85%), de 600 m > 500 m > 450 m, con recuperaciones ligeramente incompletas (3-4-5 repeticiones).
Esta “base” (velocidad del cuatrocentista -150m- y adaptación sin traumas “por arriba”) nos permitirá en un planteamiento elemental atender “el centro” y final de la carrera: de los 250 m/300 m a los 350 m/400 m. Es cierto, y se da por hecha, su atención, pero hay que tener en cuenta que cuando preparemos en el “especial”, la fase final, estaremos también entrenando la fase inicial y sobre todo la intermedia (imprescindible obsesionarse con un aceptable reparto del esfuerzo, cronometrando fracciones de 50 m o, al menos, la mitad de la distancia a recorrer).
Y, también, saber que cuando se realizan por ejemplo: 3 x 300 m al 90% con 15’ de recuperación, en la 2ª y 3ª repetición ya estamos en situaciones de generación y resistencia al lactato superiores claramente a las de la repetición 1ª y muy cerca de las sensaciones finales de un 400m.
Imagen de cabecera: Laura Fernández disputando una prueba de 400m en el Estadio Corona de Aragón. (Foto: Víctor Vieytes – El Ojo Azul)



